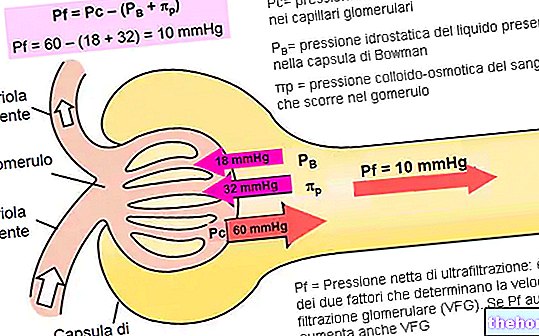
PŘIDRUŽENÍ (NEBO „KONCATENACE“ NEBO „ODKAZ“)
Až dosud jsme ošetřovali křížení Mendelian dihybrid (nebo polyhybrid) za předpokladu, že různé alelické páry se ve skutečnosti nacházejí na různých homologních chromozomových párech. Ale počet chromozomálních párů, přestože se liší druh od druhu, se liší v úzkých mezích (několik druhů) dosáhnout kolem stovky chromozomů), přičemž počet genů lze počítat na desítky tisíc.
Že postavy, které si Mendel vybral pro své experimenty, oddělovaly všechny nezávisle (aniž by si tím pletly výpočty v distribuci fenotypů v F2 polyhybridního kříže), to byla šťastná šance. Pokud byly na sousedních lokusech nalezeny dva páry alel, zákon, který bude následovat, bude se nazývat asociační zákon.
S vědomím, že velmi mnoho znaků má svůj lokus na jediném chromozomu a že jsou chromozomálními páry, které se při meióze nezávisle oddělují, lze dobře pochopit, jak často se stává, že dva páry znaků, pokud byly spojeny na chromozomu v rodičovské organismu, zůstávají sdruženi. stejně tak v gametě a tedy v organismu, do kterého přinese vlastní genetický materiál.
Vidíme tedy, že „asociace představuje“ výjimku, zdaleka ne zřídka, nezávislosti vyjádřené ve třetím Mendelově zákoně.
VÝMĚNA NEBO „PŘEKROČENÍ“ A RECOMBINACE
Když mluvíme o meióze, naznačili jsme, že existují dva různé momenty míchání genetického materiálu: jedním je segregace chromozomů v gametách a ten je pozorován Mendelem.
Druhý okamžik, který ve skutečnosti předchází, je ten, ve kterém si čtyři chromatidy z každého páru homologních chromozomů vzájemně vyměňují identické znaky. Po této výměně budou dva faktory, které byly spojeny na stejném chromozomu, místo toho v gametách nezávislé. že k výměně dochází, je úměrné první aproximaci délce chromozomu a v delších chromozomech může být výměny ještě více než.
Tento jev lze detekovat cytologicky pozorováním dostatečného počtu meióz pod mikroskopem.
Míra opětovné shody je míra, ve které se jakékoli dva znaky, které byly přidruženy v rodičovské generaci, rekombinují odlišně ve F2.
Pokud jsou dva lokusy naprosto souvislé, pravděpodobnost, že je rozdělí chiasm, bude prakticky nulová. Rychlost rekombinace bude: žádné rekombinanty. Pokud jsou dva lokusy na dvou různých chromozomech, bude rychlost rekombinace 0,5 (stejná pravděpodobnost, že se dva znaky, které byly spojeny v generaci P, náhodně ocitnou společně ve F2). Rychlost rekombinace se proto může pohybovat mezi 0,0 a 0,5. Pro malé vzdálenosti na chromozomu jsou vzdálenost a rychlost rekombinace přímo úměrné. Na delší vzdálenosti existuje možnost, že mezi dvěma lokusy proběhnou dvě výměny. Nyní bude zřejmé, že dva faktory oddělené dvěma burzami jsou opět spojeny. v tomto bodě je zřejmé, že proporcionalita mezi vzdáleností lokusů a pravděpodobností rekombinace je ztracena.
Lokusy nalezené asociované na stejném chromozomu představují „asociační skupiny“. Velmi vzdálená místa mohou mít takovou pravděpodobnost oddělení výměnou, že se chovají jako nezávislí, ale každý z nich bude spojen s nižší rychlostí rekombinace s mezilehlými lokusy.
Když jsou známy rychlosti rekombinace mezi mnoha páry genů v asociační skupině, může začít stavba „genetických map“. Je třeba mít na paměti, že vzdálenost mezi dvěma geny (a a b) je vyjádřena rychlostí rekombinace a že vzdálenost a od třetího genu c může být buď součet, nebo rozdíl vzhledem k jeho vzdálenosti od b, je možné rekonstruovat mapu vzájemných vzdáleností, což bude genetická mapa v rámci této skupiny asociací, tj. toho chromozomu.
Nyní musíme obecně uvažovat o některých konceptech, které omezují fenotypový projev genotypových znaků.
Nejprve si promluvíme o pojmech penetrace a expresivity a poté budeme věnovat zvláštní pozornost fenoménům regulace působení genu.
PENETRANCE
Penetrace genu představuje jeho schopnost projevit se ve fenotypu. Penetrance se měří statisticky počítáním frekvence fenotypů, které ukazují tento znak, ze 100 genotypů, které jej obsahují. Znak s penetrací 0,7 je znak, který se vyskytuje fenotypicky v 70% jeho genotypové frekvence.
EXPRESIVITA
Expresivita je kvantitativní hodnocení stupně fenotypové manifestace.
REGULACE GENOVÉ AKCE
Buňky produkují všechny své enzymy a proteiny stejnou rychlostí a současně. Buňky Escherichia coli mohou být například zásobovány energií a atomy uhlíku z disacharidu laktózy, protože jsou schopné je štěpit na glukózu a galaktózu díky enzymu beta-galaktosidázy. V normální E. coli, která může mít laktózu, existuje přibližně 3 000 molekul beta-galaktosidázy, což se rovná 3% proteinů této buňky; v nepřítomnosti laktózy bude na bakteriální buňku pouze jedna molekula beta-galaktosidázy. Galaktosidáza bude syntetizována z nových molekul mRNA, pokud ji lze použít. Mutantní kmeny E. coli bohaté na enzym jsou známy, i když chybí laktóza: tyto mutanty jsou ve srovnání s normálními buňkami nevýhodné, protože jsou nuceny zbytečně spotřebovávat energii a materiály k produkci enzymu, který zůstane bez substrátu. Látky, které způsobují zvýšení množství enzymu, jako je tomu u laktózy, se budou nazývat induktory, zatímco o enzymech se bude říkat, že jsou indukovatelné. Jiné látky indukují, také tyto specifickým způsobem, produkci určitých enzymů. Také například v E. coli, která je schopna vybudovat všechny své aminokyseliny s uhlíkem a amoniem (NH3), přítomnost konkrétní aminokyseliny (například histidinu) v kultivačním médiu blokuje produkci všech enzymů spojených s biosyntéza samotné aminokyseliny: o těchto enzymech bude řečeno, že jsou represivní. V bakteriálních buňkách jsou molekuly mRNA zničeny krátce po jejich vzniku, a proto řízení produkce mRNA znamená ovládání enzymatické syntézy současně čas.
OPERON
Aby vysvětlili, jak je bakteriální buňka schopná ovládat vlastní produkci enzymů, Jacob a Monod formulovali hypotézu operonu; operon je tvořen několika geny, které jsou funkčně příbuzné a zarovnané bez nespojitosti podél úseku DNA. Operon se skládá ze tří různých typů genů: promotoru, kde začíná tvorba mRNA; operátor, kde je vykonávána kontrola; jeden nebo více strukturních genů, které kódují enzymy nebo jiné proteiny. V systému beta-galaktosidázy obsahuje operon kromě systému pro beta-galaktosidázu také dva další strukturní geny kódující další enzymy podílející se na metabolismu laktózy. Tyto geny navzájem sousedí a jsou transkribovány jeden po druhém podél stejné šroubovice DNA do jediné molekuly mRNA. Takto vyrobené molekuly mRNA jsou aktivní velmi krátkou dobu, poté jsou zničeny specifickými enzymy.
Aktivita operonu je zase řízena jiným genem, regulátorem, který může být také vzdálen od operonu: tento regulátor kóduje protein, nazývaný represor, který se zdá, že se váže na DNA v operátorovém genu. mezi promotorem a strukturálními geny ve skutečnosti blokuje produkci mRNA.
Represor je zase řízen a ovládání je prováděno pomocí „signální“ látky. V případě indukovatelných enzymů je tato látka "induktorem. Induktor" se váže na molekulu represoru a upravuje svůj tvar tak, že se již nemůže přizpůsobit DNA: v tomto případě, protože mezi promotorem a strukturními geny neexistuje represor , represor může tvořit molekuly mRNA a z nich molekuly bílkovin. Po vyčerpání dodávky induktoru opět regulátor získá kontrolu, což zastaví produkci nové mRNA, tedy nových proteinů. V systému beta-galaktosidázy je induktorem laktóza nebo látka velmi podobná tomuto. Derivát: se připojí k represoru a deaktivuje jej, aby umožnil biosyntézu enzymů. V případě potlačitelných enzymů látka, která působí jako „signál“, působí jako korepresor: represor je aktivní pouze v kombinaci s korepresorem. V histidinovém systému, který zahrnuje tucet různých enzymů, je to tato aminokyselina v kombinaci se svou tRNA, korepresorem, histidinem.
ALLOSTERICKÉ INTERAKCE
Allosterické interakce, zahrnující inaktivaci enzymu změnou jeho tvaru, poskytují jiný způsob regulace metabolické aktivity buňky. Alosterické interakce umožňují přesnější řízení než systém induktor -represor operonu, ale nedosahují užitečného výsledku vyloučením biosyntézy dané látky z prvního stupně - produkce mRNA.
KONTROLNÍ SYSTÉMY V EUKARIU
Existuje několik faktů, které vedou k domněnce, že mezi rostlinami a živočichy funguje a dominuje systém regulace podobný operonu. Chromozomy těchto organismů se výrazně liší od chromozomů E. coli a jiných prokaryot. Kontrola genů v těchto buňky jsou velmi odlišné.Mechanismus mitózy je takový, že každá buňka dané rostliny nebo zvířete vlastní všechny informace
genetika přítomná v oplodněném vajíčku. Proto většina genů v jakékoli specializované buňce zůstane po celý život buňky neúčinná. DNA v těchto buňkách je vždy spojena s proteiny. Je tedy možné, že genová represe v eukaryotech vyžaduje právě tuto asociaci mezi DNA a proteiny.